|
|
|
|
 A Company of Union of Scientists in Bulgaria
|
|
|
Abstract The first two enzymes (phenol 2-monooxygenase (EC 1.14.13.7) and catechol 1,2- dioxygenase (EC 1.13.11.1)) of phenol biodegradation pathway in Tr.cutaneum were investigated. The cultivation of the strain was performed in two different nutrient media -–complete and minimal salt media. The enzyme activities of crude cell extracts from both types of media were determined. A comparison between our data and the reported in the literature was made. The results reported confirm the potential of the investigated strain to be used in technologies for microbial detoxification of phenol polluted industrial waste water. Key words:Trichosporon cutaneum,phenol degradation,phenol hydroxylase,catechol 1,2 dioxygenase
Degrading, biotransformation and utilization of xenobiotics are between the most interesting and important topics of modern ecology. Typical representatives of this group of environmental pollulants are phenol and its derivatives. Unsubstituted phenol, which is metabolized by the 3-oxoadipate pathway enters the central metabolism through succinate and acetil-S-CoA. The 3-oxoadipate pathway involves six enzymes, which all are located in the cytosol (Fig1) [1]. Phenolic compounds , as by-products of industrial reaction, are a serious threat to the environmrnt. Sertan soil living yeast use flavin adenin dinucleotide (FAD)-containing enzymes to metabolize these compounds through hydroxylation. The phenol hydroxylase from Trichosporon cutaneum hydroxylates phenol to catechol. Exept from phenol, which is the best substrate, the enzyme also, converts hydroxyl-, amino-, halogen- or methyl-substituted phenols [2]. Catechol 1,2 dioxygenase, the second enzyme of phenol metabolism is the ring-cleaving enzyme which catalysis the process of catechol oxidation. This enzyme is induced simultaneously with the first enzyme (phenol 2-monooxygenase) and the third enzyme (muconate cycloisomerase) of phenol metabolism[3].
Fig.1. Phenol metabolism including the first steps of the ortho- fission pathway
Materials and Metods The investigated strain of Trichosporon cutaneum NBIMCC N 2414 shows well expressed capability for metabolizing high concentration phenol (up to 1g/l), utilizing it as a sole carbon and energy source for its growth and development [4]. The strain cultivation is carried out on a minimal salt medium (MM) [5] or on a complete medium (YEPD). In both cases the phenol concentration is 0.5 g/l. Intracellular enzyme levels are assayed in crude extract. Enzyme activities are determined at ambient temperature (21oC) by the following methods: phenol 2- monooxygenase by following NADPH oxidation at 340 nm [6]; catechol 1,2 dioxygenase by following the increase in absorbance at 260 nm [7]. Protein was determined by method of Lowry et al. [8]. One Enzyme unit is defined as amount of enzyme which catalyzes the disappearance of 1 m mol substrate, cosubstrate or the formation of 1 m mol product per minute per mg protein. Results and Discussion
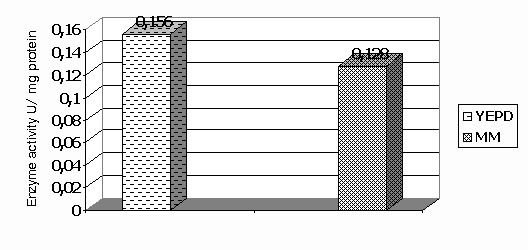
In the present study, the specific activities of the first two enzymes involved directly in phenol degradation - phenol hydroxylase and catechol 1,2–dioxigenase were investigated (Fig 2 and Fig 3). Figure 2. Influence of growth medium on the specific enzyme activity of phenol hydroxylase
Figure 3. Influence of growth medium on the specific enzyme activity of catechol1,2 dioxygenase Cell growth on YEPD with phenol showed higher specific activity of investigated enzymes then cells grown on minimal medium. We suggest, that the complete medium ensure better vitamin balance and other favorable conditions for development of the strain in the presence of phenol. The activities of crude extracts of cell grown on glucose containing media were too low to be measured directly.The phenol hydroxylase and catechol 1,2 dioxygenase isolated by usfrom Trichosporon cutaneum exhibit certain similarities and differences in comparison with similar enzymes isolated from the other microorganisms (Table 1). We should point out, that the data shown on this table might differ in their values not only because the investigated strains but the cultivation conditions, as well, are not exactly the same. Our results show a phenol hydroxylase activity higher than the other cited data. The values for the second enzyme, catechol 1,2 dioxygenase vary significantly. That fact may be explained with the less substrate specificity, typical for this enzyme [13]. Table1. Specific enzyme activity of phenol hydroxilase and catechol 1,2 dioxygenase from different microorganisms.
On the basis of our previous and recent analyses, we consider that the investigated strain has good perspectives for its application in protecting and remediating the environment from phenol pollutions. References 1. Spanning A., H. Neujahr, FEMS Microbiol. Lett., 1991, 77, 163-168. 2. Enroth C., H. Neujahr, G. Schnaider, Y. Linqvist, Structure, 1998, 6, 605-617. 3. Spanning A., H. Neujahr, J. Gen. Microbiol., 1990,136. 1491-1495. 4. Shivarova N., A. Christov, B. Atanasov, N. Peneva, N. Ivanova, Z. Alexieva, 1996, “ Ecology 96”, Burgas. 5. Godjevargova T., Z. Alexieva, D. Ivanova, N. Shivarova, Process Biochem., 1998, 33, 831-835. 6. Neujahr H., A. Gaal, Eur. J. Biochem., 1973, 35, 386-400. 7. Varga J., H. Neujahr, Eur. J. Biochem., 1970, 12, 427-434. 8. Lowry O., N. Rosenbrought, A. Farr, R. Randall, J. Biol. Chem, 1951, 193, 265. 9. Gaal A., H. Neujahr, Arch. Microbiol., 1981, 130, 54-58. 10. Chang S, C. Li, S. Hiang, M. Chang, Appl. Microbiol. Biotechnol., 1995, 43, 534-538. 11. Hinteregger Ch., R. Leitner, M. Loidl, A. Fershl, F. Streichsbier, Appl. Microbiol. Biotechnol., 1992, 37, 252-259. 12. Topalova Y., R. Dimkov, D. Merjanov, Biotechnol&Biotechnol. Eq., 1995, 9, 55-59. 13. Neujahr H., J. Varga, Eur. J. Biochem., 1970, 13, 37-44. |
|